Antibodies
to gliadin, a component of gluten, have frequently been reported in schizophrenia.
Highly immunogenic B cell epitopes along its length are homologous to
numerous proteins relevant to schizophrenia, including members of the
DISC1 interactome, glutamate, dopamine and neuregulin signalling networks,
and plasticity or myelination pathways. Antibodies to gliadin may cross
react with these key proteins, as has already been observed with synapsin
1 and calreticulin. Gliadin may thus be a causative agent in schizophrenia,
under certain genetic and immunological conditions, producing its effects
via antibody mediated knockdown of multiple proteins relevant to the
disease process. Because of such homology, an autoimmune response may
be sustained by the human antigens that resemble gliadin itself, a scenario
supported by many reports of immune activation both in the brain and
in lymphocytes in schizophrenia. Gluten free diets and removal of such
antibodies may be of therapeutic benefit in certain cases of schizophrenia.
Introduction
A number
of studies from China, Norway, and the USA have reported the presence
of gliadin antibodies in schizophrenia 1-5. Gliadin is a component of gluten,
intolerance to which is implicated in coeliac disease 6. Both gluten intolerance and coeliac
disease have also been associated with schizophrenia 7, 8, and remission of schizophrenia, in
specific subsets of patients , has occasionally been reported following
the instigation of a gluten-free diet 9, 10. Gliadin is a polyglutamine repeat
protein (Fig 1), and de facto, a homologue of the mutant polyglutamine
proteins in Huntingtons disease, Dentatorubropallidoluysian atrophy (DRPLA)
, spinal and Bulbar Muscular Atrophy (Kennedy disease) and Spinocerebellar
ataxias 11. Gliadin antibodies have also been
found in Huntingtons disease and spinocerebellar ataxias and gluten per
se has been associated with various forms of ataxia 1213. These studies may implicate gliadin
in the pathology of these diseases. This is not the subject of this article.
As reported
below, polyglutamine repeats are highly immunogenic, the more so with each
addition of glutamine. A number of schizophrenia susceptibility gene products
contain polyglutamine repeats, while others also display a high degree
of homology to other regions of gliadin. Gliadin antibodies may thus play
a role in the pathology of schizophrenia by cross-reactive targeting of
key schizophrenia-related proteins.
Results
Gliadin is
a polyglutamine (polyQ) repeat protein with an internal contiguous sequence
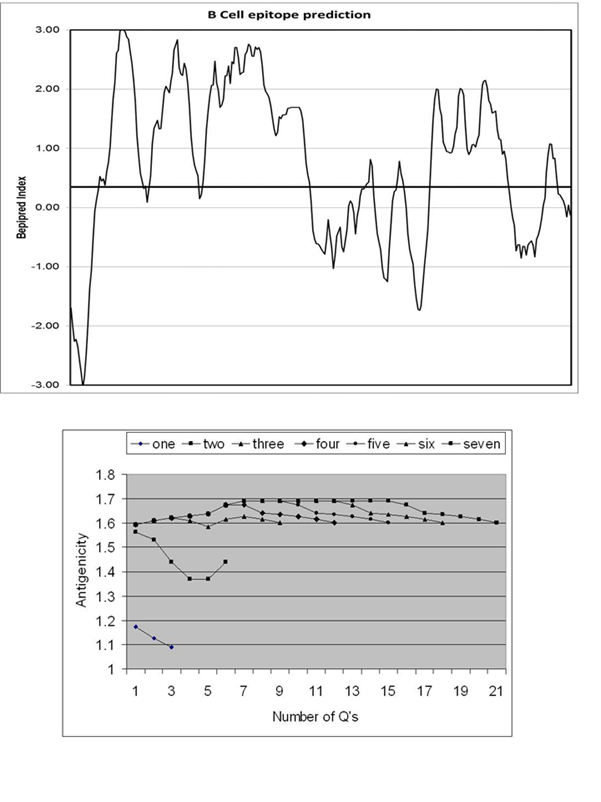
of 22 glutamines (Fig 1). Gliadin is highly immunogenic and 181/296 (61%)
of its residues are considered as B cell epitopes with the server-defined
cut off index of 0.35 (Fig 2). http://www.polygenicpathways.co.uk/gliadin.htm.
Polyglutamine repeats are also immunogenic, and all are above the threshold
of 0.35 (Fig 2). The antigenicity increases with the number of glutamine
repeats. The BLAST of gliadin (whole protein) versus the human proteome
yielded 29 significant results, again including highly relevant proteins,
many, but not all, influenced by the polyglutamine repeat (Table 1). These
proteins belong to members of the DISC1 interactome and also include pre-
and postsynaptic proteins related to glutamate, GABA, and neuregulin signalling.
They are also involved in dopaminergic function, myelination, and dendritic
spine development and to neurogenesis, inflammation and oxidative stress
(Fig 3).
A more detailed
analysis revealed an interesting type of homology that is common to many
more proteins than are listed in Table 1. This is exemplified in Fig 4.
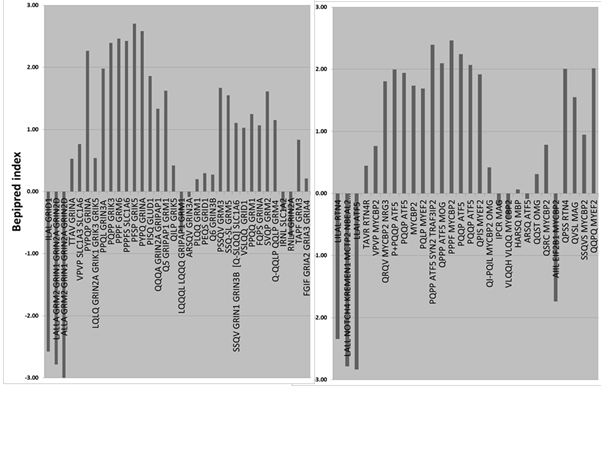
The Clustal alignment of KCNN3 with gliadin shows a non-extensive homology
with 18% amino acid identity shared by the two proteins, within the homologous
region. The gliadin protein is characterised by many short repeat motifs,
other than polyglutamines. These include (PQPQP) *4 ;(PQPQ) *5; (QPQP)*5; (PQQP)*2;
(QQPY)*2; (QPQPQ)*2; (QQQQF)*2 and (VLQQ)*2. Some of these motifs can
also be found in the KCNN3 protein, which, with the polyglutamine repeat
contains 9 such areas of identity, many concentrated in a 13 amino acid
contiguous tridecapeptide (QPQPPQLQQQQ). The overall gliadin/KCNN3 identity
including these contiguous peptides is 5.3%. However, these matching gliadin/KCNN3
motifs extend over the whole length of the gliadin protein, which displays
31% identity with these KCNN3 fragments.
These fragments
are mostly (12/15) within highly immunogenic regions of the gliadin protein.
Because this immunogenicity extends over many different regions, several
different antibodies to gliadin or to its partially digested fragments
are likely to be produced. These repeat motifs and their presence in human
proteins are likely to dramatically increase the likelihood of cross-reactivity
between gliadin, gliadin fragments and human antigens.
These areas
in human proteins were identified by sequential BLASTS of contiguous 25
amino acid fragments, along the length of the gliadin protein, each BLAST
overlapping by 5 amino acids. By plotting these homologues along the length
of the gliadin protein, these multiple repeats and their antigenicity could
be identified simultaneously. In all, a total of 459 human proteins contain
at least one of these tetrapeptide matches, 60 with pentapeptide matches,
and others with longer contiguous matches as shown in Table 2.
This procedure
allowed the definition of the number of gliadin matches within each human
protein, and of the antigenic index of each of these matches. These results,
for 5 or more matches are shown in Table 3: FOXP2, SMARCA2, NUMBL, KCNN3,
and RAI1 contain from 14 to 22 of such matches. Certain key gene products
including DISC1, neuregulin 2, dysbindin and synapsin 2 contain 5 or more
of these gliadin consensus sequences. In fact, of the 459 proteins identified,
158 (34%) are the products of genes listed as susceptibility candidates
in association studies. (See http://www.polygenicpathways.co.uk/gliadin.htm for
details) An example of peptide matching to the antigenicity profile is
shown in Fig 5 for KCNN3. This also allowed antigenicity mapping by family
as shown in for glutamate related proteins or for myelin related proteins,
many of which match gliadin in highly immunogenic regions (Fig 6).
A number
of autoantibodies have been reported in schizophrenia. Their targets include
the dopamine (DRD2) 14, glutamate (GRIN1) 15, acetylcholine (CHRNA7) 16 and opioid (OPRM1) receptors 17 , heat shock protein 60 (HSPD1) 18 and hsp90 (HSP90A1) 19, MYC binding protein 2 (MYCBP2) 18 nerve growth factor (NGF) 20 and striatin (STRN) 21 all of which are homologous to gliadin,
with MYCBP2 and striatin particularly well represented (Fig 7). Myc inhibits
myelination, and is also involved in dendrite and synapse formation 22, 23. Striatin plays an important role in
dendritic spine development 24.
Discussion
The high
immunogenicity of gliadin, over almost its entire length, suggests that
multiple antibodies could be formed by presentation of diverse antigens
in different cellular and tissue compartments. Such antibodies might be
produced to the entire protein, or to its partially digested fragments.
The homology with key schizophrenia related proteins, often covering many
regions of gliadin is striking and suggests that gliadin antibodies could
also target these human proteins. Indeed 9 of the autoantigens reported
in schizophrenia patients are homologous to gliadin. It has also been shown
that gliadin antibodies cross react with calreticulin and synapsin 1 25, 26. Synapsin 1 was homologous to the particular
gliadin tested (PQQQP, PQQP and PLQQ) while calreticulin was not. It was
however homologous to gamma gliadins from Triticum aestivum and Triticum
urartu (VPRD), Triticum monococcum (VRPD) and a related goat grass, Aegilops
searsii (PVIQ).
These homologous
sequences are short, and in most cases, tetrapeptides, although higher
degrees of homology were observed in many proteins (from 5 to 9 amino acids,
not including polyglutamine repeats (Table 2). Antibodies are quite capable
of binding to such short epitopes 27. In addition the repeat motifs in gliadin
have already been noted and are, per se, immunogenic 28.
The key pathological
features of schizophrenia include reduced dendritic spine density and synaptic
poverty 29, deficits in myelination and oligodendrocyte
cell loss 30, 31, impaired neuregulin signalling 32and imbalances in glutamate and dopamine
neurotransmission 33. Many gene products covering these
networks are homologous to gliadin. The DISC1 network, connected to many
of these areas34 is also clearly targeted by gliadin
homologues (Fig 1).
Antibodies
are able to enter the brain via blood-brain barrier transporters 35 and can also enter cells via a high
affinity immunoglobulin receptor, tripartite motif-containing 21 (TRIM21) 36.
This suggests
that antibody related protein knockdown, of multiple proteins relevant
to schizophrenia could be a direct consequence of gliadin allergy. Many
studies have reported evidence of immune activation in schizophrenia patients,
both in the brain 37, 38 and in lymphocytes 39, 40, and autoimmune attack of certain cells
may well explain some of the ongoing pathology of schizophrenia, for example
oligodendrocyte 30 and grey matter loss 41.
Following
digestion, gliadin will be broken into peptide fragments that may also
find their way into the brain via the circulation, and uptake via peptide
transporters. As partial homologues of many relevant proteins, they may
also be able to interfere with the signalling processes controlled by these
proteins. Several pharmacological effects of gliadin or gluten have indeed
been noted. For example gluten peptides have opioid activity 42: Gliadin peptides are also able to
activate protein kinase A 43 and bind to the chemokine receptor
CXCR3 44. They are also able to interfere with
epidermal growth factor signalling in a number of cell lines 45 and activate nuclear factor kappa beta
signalling in monocytes 46 . This type of homology with human
proteins may apply to allergens in general, whose deleterious effects are
not necessarily restricted to immune activation, but also to the possibility
of interaction with a multitude of host proteins that they resemble.
Similar protein
matches are found in many proteins expressed by the viruses implicated
in Alzheimers disease 47 or schizophrenia, and indeed these
viral consensus sequences tend to be longer. For example hexapeptide identity
between influenza viral proteins and several schizophrenia relevant proteins
has been noted: These include reelin, neurexin 1-alpha and DISC1 48. In fact several hundred schizophrenia
susceptibility gene products display this type of homology to diverse viruses
and parasites (T.Gondii and B. Burgdorferri) implicated as risk factors
in schizophrenia. It has recently been shown that DNA from many common
non-retroviral viruses is integrated into mammalian genomes 49. BLAST analyses of the human proteome
also shows that thousands of human proteins contain these viral (or in
this case allergen) contiguous matching sequences. Indeed this type of
viral homology appears to cover the entire human genome. (See http://www.polygenicpathways.co.uk/blasts.htm ).
The human proteome has been estimated to contain ~33869 proteins with an
average length of 375 amino acids 50. For pentapeptide matches, this yields
a figure of 370*33869 potential matching blocks (12.53 million). These
building blocks are identical to those in viral, bacterial, fungal and
allergen proteins. Upon infection or ingestion, these pathogenic proteins
are likely to seed havoc in the panoply of the hosts signalling networks
via the mechanisms described above, and are likely to contribute to the
pathology of many human diseases.
It should
also be noted that the viral and allergen protein homology is of course
reflected at the DNA level and that viral double stranded DNA, plant or
bacterial DNA is indistinguishable from our own. It is thus plausible that
many gene association studies, using blood samples, have been indexing
infection and ingestion as well as identifying key susceptibility genes.
This is less of a problem when using long DNA probes and in no way detracts
from the gene association results, whose relevance is generally supported
by a plethora of experimental data related to the function of the genes
identified. However, the confluence of gene and risk factor homology suggests
that many genes are risk factors precisely because they encode for proteins
that are homologous to those expressed by the viral, bacterial and allergen
risk factors. However, they may only act as risk factors when such confluence
is achieved, perhaps explaining the problems of replication in both gene
and risk factor association studies.
Clearly gliadin,
a major dietary component, cannot cause schizophrenia in all cases. Gluten
intolerance and gliadin allergy are evidently related to the immune system.
Many immune related susceptibility genes (few of which were encountered
in this study) have been reported in association studies, including genome-wide
association studies 51. The high proportion of other types
of schizophrenia susceptibility gene products related to gliadin suggests
that those genes that encode for proteins with gliadin homology may be
considered as risk factors if and when their products are homologous to
a particular form of gliadin. Many different forms of gliadin, from diverse
plant and bacterial species exist, as do many polymorphic genes, and their
resultant differing protein sequences. The marriage of genes and risk factors,
and the status of our immune system are thus three variables whose convergence
may be obligatory to initiate the processes described above.
Gliadin antibodies
and gluten intolerance have often been associated with schizophrenia, and
in some cases a gluten free diet has been reported to evoke the remission
of symptoms (see introduction). These data suggest that this is more than
a simple association and that gliadin antibodies could well be the causative
agent of schizophrenia in genetically and immunologically compromised individuals,
and illustrate how this might be achieved. If so, then the instigation
of a gluten free diet, as already shown 9, 10, may be an effective substitute for
drug related interventions. However in certain cases, even if gliadin is
removed, the homologous human proteins might well be able to sustain the
production of further antibodies, due to the permanence of autoantigens
in the human biological network. Antigen and antibody removal by immunoadsorption
techniques, or immunosuppression, might thus prove to be effective therapies.
Ways of identifying the subsets of patients, who might benefit from such
strategies, including routine antibody detection, may have a marked effect
on the prevalence and severity of schizophrenia.
Acknowledgements:
I would like to thank the many authors who have provided reprints and encouragement.
Methods.
Gliadin is
a polyglutamine (polyQ) repeat protein with an internal stretch of 22 glutamines.
This sequence as well as gliadin or gliadin internal fragments were screened
against the human proteome using the filters schizophrenia, glutamate,
dopamine or myelin to trawl for proteins that might be related to gliadin
(BlastP)52. Without these filters, many other
polyglutamine proteins (Huntingtin, ataxins, the androgen receptor etc.)
masked any underlying results. B-Cell epitopes within the gliadin protein
were identified using the BepiPred server http://www.cbs.dtu.dk/services/BepiPred/,
which predicts antigenicity related to the charge and hydrophobicity properties
of the peptide 53. The BLAST results and supplementary
data can be visualised at http://www.polygenicpathways.co.uk/gliadin.htm where
a NextBio highlighting tool provides details for all gene symbol abbreviations.
Table
1
Polyglutamine
repeat proteins or proteins with significant overall homology to gliadin
(BlastP gliadin vs. schizophrenia, glutamate, dopamine or myelin).
Brief descriptions of function are provided for each protein. The number
of glutamines is represented by, for example, Q22, or by the Q containing
sequence in the human protein. E values are provided for the BLAST: If
none is shown, the results were not significant (P> 0.05)
Presynaptic
protein regulating neurotransmitter release 59
GRIPAP1
| GRIP1 associated protein 1
QQQQQEQEEALKQ-
Part
of the postsynaptic scaffold for glutamate (AMPA) receptors 60
MAP1A
: microtubule-associated protein 1A
QQTHEQQQQ
Part
of the NMDA receptor postsynaptic density, that controls dendrite
branching and synapse formation 61; Binds to postsynaptic density proteins
that connect to NMDA Kainate , AMPA and ERBB receptors
Part
of the postsynaptic density that regulates glutamate receptor signalling 62
RORB
: RAR-related orphan receptor B
QKHQQRLQEQRQQQ
Little
is known about this protein: However it binds to MAP6, a microtubule
protein that controls synaptic organisation, in particular of glutamatergic
synapses where it controls the expression of the glutamate transporter
and presynaptic genes , synaptophysin and GAP-43 , spinophilin
and MAP2 63.
Regulates
the development of neurones, astrocytes and oligodendrocytes 64
DGCR6
: DiGeorge syndrome critical region gene 6
QKHQEAQQACRPHNLPVLQAAQQ
BLAST E value 0.001
Little
is known except that is binds to the GABA-b receptor GABBR1 65
SP4
: Sp4 transcription factor
QNAQDQSNSLQQVQIVGQPILQQIQIQQPQQQ
BLAST E = 9e-05
Regulatesglutamate
(GRIN2A), GABA (GABRA4) receptors (and many others): Also controls
dendritic function 66-68
Dopamine and serotonin related
FOXP2
Forkhead box P2
Q22
BLAST 3e-25
Controls
pathways involved in embryonic and nervous-system development,
neurogenesis, cell migration and cell death. FOXP2 knockout mice
have lower cerebral dopamine concentrations. Expressed in dopamine
and cyclic adenosine 3',5'-monophosphate-regulated phosphoprotein
containing neurones in the cerebral cortex FOXP2 also controls
dendritic spine development and synaptic plasticity. 69-72
LMX1A
and B control the development of midbrain dopamine neurones 73
LMX1B
: LIM homeobox transcription factor 1, beta
HQQQQE QQ
KCNN3
potassium intermediate/small conductance calcium-activated channel,
subfamily N, member 3
Q19
BLAST
E Value: 2e-22
Regulates
the activity of dopamine and serotonin neurones and plays a role
in glutamate-mediated long term potentiation 74, 75
MAGEL2
MAGE-like protein 2
QQAQASGPQ
BLAST
E Value 2e-05
MAGEL2
and necdin bind to FEZ1 and are involved in the control of axonal
growth and in the development of brain dopamine, serotonin and
noradrenaline neurones 76
NALCN
: sodium leak channel, non-selective
QQQSCSIIHSLRESQQQE
Sodium
channel activated by reducing extracellular calcium levels: It
is activated by substance P and neorotensin in ventral tegmental
and hippocampal neurones 77
PDCD6IP
programmed cell death 6 interacting protein
E
value 6e-06
Apoptosis
inhibitor that also binds to D1 and D3 dopamine receptors 78
POU3F2
POU class 3 homeobox 2
brain-specific
homeobox/POU domain protein 2
Q18
E
value = 4e-15
Controls
the expression of neural progenitors and of the tryptophan hydroxylase
gene (TPH2) 79, 80
PPP1R3F
: protein phosphatase 1, regulatory (inhibitor) subunit 3F
QQQQLPQLEPQ
Protein
phosphatase 1 regulates the activity of many transmitter systems
(glutamate, GABA, dopamine . serotonin, inter alia ): It also modifies
dendritic spine development . 81-86
RANBP9
| RAN binding protein 9
Q8
Binds
to the D1 dopamine receptor as well as to nerve growth factor receptors
NGFR and trkB (NTRK2) , androgen and glucocorticoid receptors and
DISC1 87-91
SMARCA2
: SWI/SNF related, matrix associated, actin dependent regulator
of chromatin, subfamily a, member 2
Q2
BLAST
E value 3e-15
Binds
to the promoter of the noradrenaline transporter SLC6A2 : It also
controls neuronal development on the mid hindbrain 92, 93
SMARCC1
: SWI/SNF related, matrix associated, actin dependent regulator
of chromatin, subfamily c, member 1
QQMEQQQHGQNPQQAHQH
And
EQQRQQLLTERQNFHMEQLKYAELRARQQMEQQQHGQNPQQ
Regulates
the function of the glucocorticoid receptors (NR3C1), and Nurr77
(NR4A1) 94 both of which play an important role
in the control of dopamine neurones 95
Neuregulin , NOTCH and growth
NRG2
: neuregulin 2
QQQQQQREEQQQQQQQQRERQQQQEQQQ
This
particular neuregulin is localised to dendrites and controls the
expression of DISC1 ; It also controls free radical release from
microglia 96-98
NUMBL
numb homolog (Drosophila)-like
Q20
BLAST
E value 6e-13
Important
regulator of cortical neurogenesis 99, 100
RAI1
retinoic acid induced 1
Q14
BLAST
E Value = 4e-10
Regulates
BDNF expression and also regulates autoimmunity by inhibiting lymphocyte
activation and and triggen receptor signalling 101, 102
Dendritic spines and plasticity
HDAC4
: histone deacetylase 4
QQQHQQFLEKHKQQFQQQQLQ
BLAST
E value 0.006
Histone
deacetylases regulate gene transcription by modifying histones.
HDAC4 is localised in the brain and shuttles between the nucleus
and cytoplasm in response to neuronal activity. HDAC4 is also localised
in dendritic spines associated with the postsynaptic density: HDAC9
also shuttles from nucleus to cytoplasm in response to neuronal
activity, and controls dendritic spine density 103-105
HDAC5
| histone deacetylase 5
QQQEMLAAKQQQEMLAAKRQQELEQQRQREQQRQEELEKQRLEQQ
BLAST
E value 5e-05
HDAC9
: histone deacetylase 9
QQQQQIQKQLLIAEFQKQHENLTRQHQAQLQEHIKLQQELLAIKQQQ
MCF2
: MCF.2 cell line derived transforming sequence
QQDQLTERDKFQISLQQNDEKQQ
Controls
dendritic spine development 106
MED25
| mediator complex subunit 25
VQQQ
BLAST
E Value 0.001
Regulates
the retinoid receptor RAR , which controls dendritic spine formation, 107
NR4A3
: nuclear receptor subfamily 4, group A, member 3 also known as
nor-1
QQQHQQ
Regulates
pyramidal neurone survival and hippocampal axon guidance and controls
neurite extension 108, 109
PURA
: purine-rich element binding protein A
EQLHQQQQQQQEE
BLAST E value 0.014
A
single stranded DNA (e.g. viral) binding protein involved in the
transport of RNA to dendrites. Plays and important role in postnatal
brain development and also controls the transcription of myelin
basic protein in oligodendrocytes 110-112
Myelin related
FOXJ3
| forkhead box J3
QQQP
BLAST
E value 5E-0.5
Involved
in oligodendrocyte proliferation 113
NCOR2
| nuclear receptor corepressor 2
Q17
BLAST
E value = 6E-11
Controls
thyroid hormone, retinoic acid and steroid receptors114. Retinoids play a key role in dopamine
function, dendritic spine development and oligodendrocyte differentiation 115-117
PEG10
| paternally expressed 10
No
Polyglutamines
BLAST E value 0.006
Controls
the expression of myelin basic protein 118
SGCE
: sarcoglycan, epsilon
QTQQNLPHQTQIPQQQ
Sarcoglycans
are typically associated with muscle cells: However SGCE is expressed
in Schwann cells, and in monoaminergic neurones and is believed
to regulate dopaminergic activity 119-121
Development and neurogenesis
DISC1
: disrupted in schizophrenia 1
QQLRREIEEQEQQLQ
Key
hub gene connected to many schizophrenia networks 34
MACF1
: microtubule-actin crosslinking factor 1
QLQQLQSQLAHQTEQKTLQKQQNTCHQQ
MACF1
knockdown impairs cortical and other brain area development :
It also plays a role in growth cone advance 122, 123
PCM1
: pericentriolar material 1
QQQQRELKQLQEE
Involved
in neuronal migration and neurogenesis 124
PCNT
: pericentrin
QQQALHSQQQ
Regulates
the neuronal progenitor cells in the developing cortex 125
SEL1L3
: sel-1 suppressor of lin-12-like 3
QQQQQ
Little
is known, except that SEL1L3 binds to tubulin TUBB3 which is involved
in cortical development and neuronal migration 126
Endocytosis
CHERP
| calcium homeostasis endoplasmic reticulum protein
Q12
BLAST
E value 7e-08
No
data: Endoplasmic reticulum location
CLINT1
: clathrin interactor 1
QQQNMQQ
Involved
in clathrin mediated endocytosis and the endosomal transport of
SNARE proteins including STX7 127
STX7
: syntaxin 7
QQLQQKQQ
Involved
in endosomal to lysosomal traffic and in phagocytosis. Also expressed
in neutrophils and macrophages 128-131
Apoptosis and oxidative stress
EP400
| E1A binding protein p400
Q22
BLAST
E value 1E-16
Involved
in the regulation of oxidative stress 132
HIPK3
: homeodomain interacting protein kinase 3
QQKLTSAFQQQH
Involved
in the Fas apoptosis pathway, and also controls the expression
of cytochrome p450 CYPA11, the enzyme responsible for pregnelonone
synthesis, the firsts step in steroid hormone production 133, 134
SETD2
: SET domain containing 2 (a histone methyltransferase)
QREAQKQQQQMQ
Involved
in embryonic vascular development 135
SHISA5
protein shisa-5 precursor
No
polyglutamines
BLAST E value e-04
Localised
in the endoplasmic reticulum and nuclear membrane and involved
in p53 and p73 mediated apoptosis 136-138
TFAP4
| transcription factor AP-4
E
value 0.002
AP4
controls the expression of p21 a cell cycle cyclin dependent kinase
inhibitor induced by p53 139
Immune system
HLX
| H2.0-like homeobox
Q8
BLAST
E value 6e-06
Regulates
interferon gamma production in natural killer cells 140
HOXA10
| homeobox A10
PPQQQPPPPPQP
E
value 0.012
Part
of a vitamin D3 dependent pathway regulating haematopoesis 141
Acute
phase protein involved in inflammatory processes 142
NFAT5
| nuclear factor of activated T-cells 5, tonicity-responsive
Q17
BLAST
E value 1e-16
Plays
an important role in the immune system and in water homoeostasis
: Polydipsia (excessive water drinking) and water imbalance have
been observed in schizophrenia 143, 144.
Translation initiation
ABCF1
| ATP-binding cassette, sub-family F (GCN20), member 1
Involved
in cranial bone development 66, 67, 109, 146, 147
ARID3B
| AT rich interactive domain 3B (BRIGHT-like)
Q11
BLAST
E value 1e-05
No
data
PHC2
| polyhomeotic homolog 2
VIQQQPQPQQQQPPPQQ
BLAST
E value6E-0.6
No
data
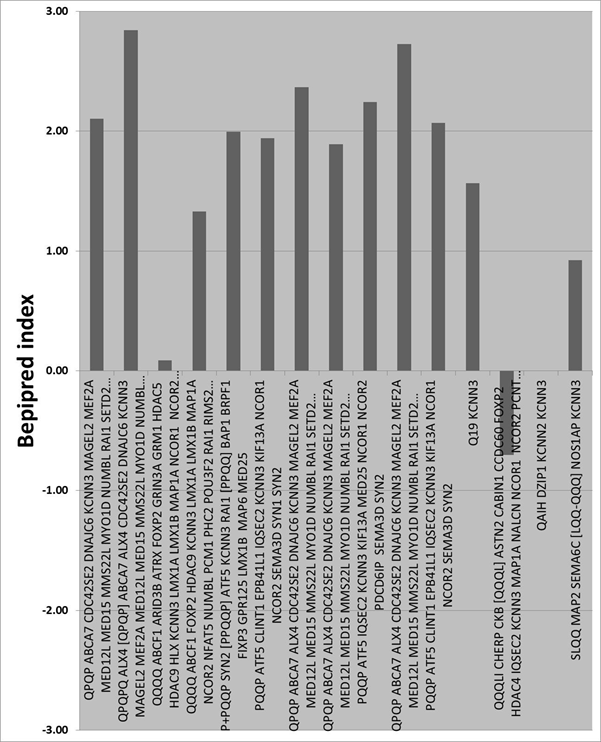
Table
2 Proteins with hexapeptide, or greater, matches to the gliadin protein.
The consensus sequence is shown on the left followed by the gene symbols.
N is the number of contiguous amino acids : DISC1 (N=5) is included for
interest: Q = glutamine, followed by the number of Qs e.g. Q22. Full
tables with abbreviations and data provided by a NextBio highlighting
service are available at http://www.polygenicpathways.co.uk/gliadin.htm
The B cell
epitope index values of the gliadin protein (amino acid 1-276) as predicted
by the Bepipred server. The cut-off value of 0.35 is illustrated by the
thick horizontal line.
Bottom
The B cell
epitope indices for various glutamine repeats (QQQ) * 1-7
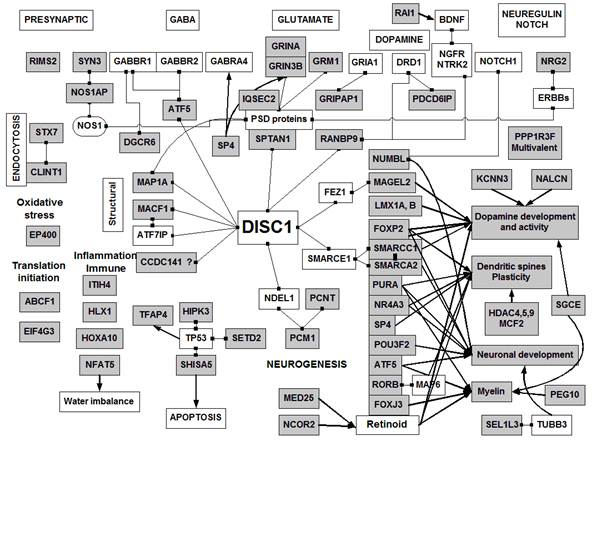
Fig 3
An interactome
network for the proteins identified in this study. Proteins in grey boxes
show a significant degree of homology with gliadin in the BLAST analyses,
or are polyglutamine repeat proteins. Linked boxes represent binding between
two components; these data were gleaned from the protein interaction section
of Entrez Gene. Arrows indicate other effects on transcription or other
functional effects. See Table 1 for details.
Fig 4
A) Clustal
line up between gliadin and the potassium channel, KCNN3
B) Gliadin
consensus sequences with the KCNN3 protein
C) KCNN3
consensus sequences within the gliadin protein
1. Jin,S.Z. et al. A Study of Circulating
Gliadin Antibodies in Schizophrenia Among a Chinese Population. Schizophr.
Bull.(2010).
2. Samaroo,D. et al. Novel immune
response to gluten in individuals with schizophrenia. Schizophr. Res.118,
248-255 (2010).
3. Cascella,N.G. et al. Prevalence
of Celiac Disease and Gluten Sensitivity in the United States Clinical
Antipsychotic Trials of Intervention Effectiveness Study Population. Schizophr.
Bull.(2009).
4. Reichelt,K.L. & Landmark,J. Specific
IgA antibody increases in schizophrenia. Biol. Psychiatry.37,
410-413 (1995).
5. Dohan,F.C., Martin,L., Grasberger,J.C.,
Boehme,D., & Cottrell,J.C. Antibodies to wheat gliadin in blood of
psychiatric patients: possible role of emotional factors. Biol. Psychiatry.5,
127-137 (1972).
6. Tjon,J.M., van Bergen,J., & Koning,F.
Celiac disease: how complicated can it get? Immunogenetics.62,
641-651 (2010).
7. Samaroo,D. et al. Novel immune
response to gluten in individuals with schizophrenia. Schizophr. Res.118,
248-255 (2010).
8. Cascella,N.G. et al. Prevalence
of Celiac Disease and Gluten Sensitivity in the United States Clinical
Antipsychotic Trials of Intervention Effectiveness Study Population. Schizophr.
Bull.(2009).
9. Kalaydjian,A.E., Eaton,W., Cascella,N., & Fasano,A.
The gluten connection: the association between schizophrenia and celiac
disease. Acta Psychiatr. Scand.113, 82-90 (2006).
10. Kraft,B.D. & Westman,E.C. Schizophrenia,
gluten, and low-carbohydrate, ketogenic diets: a case report and review
of the literature. Nutr. Metab (Lond).6, 10 (2009).
11. Robertson,A.L., Bate,M.A., Androulakis,S.G.,
Bottomley,S.P., & Buckle,A.M. PolyQ: a database describing the sequence
and domain context of polyglutamine repeats in proteins. Nucleic Acids
Res.(2010).
16. Chandley,M.J., Miller,M.N., Kwasigroch,C.N.,
Wilson,T.D., & Miller,B.E. Increased antibodies for the alpha7 subunit
of the nicotinic receptor in schizophrenia. Schizophr. Res.109,
98-101 (2009).
17. Tanaka,S. et al. Autoantibodies
against four kinds of neurotransmitter receptors in psychiatric disorders. J.
Neuroimmunol.141, 155-164 (2003).
18. Wang,X.F. et al. Studies characterizing
60 kda autoantibodies in subjects with schizophrenia. Biol. Psychiatry.53,
361-375 (2003).
19. Kim,J.J. et al. Identification
of antibodies to heat shock proteins 90 kDa and 70 kDa in patients with
schizophrenia. Schizophr. Res.52, 127-135 (2001).
20. Shcherbakova,I.V. et al. Leukocyte
elastase and autoantibodies to nerve growth factor in the acute phase
of schizophrenia and their relationship to symptomatology. World J.
Biol Psychiatry. 5, 143-148 (2004).
21. Poletaev,A.B., Morozov,S.G., Gnedenko,B.B.,
Zlunikin,V.M., & Korzhenevskey,D.A. Serum anti-S100b, anti-GFAP and
anti-NGF autoantibodies of IgG class in healthy persons and patients
with mental and neurological disorders. Autoimmunity.32,
33-38 (2000).
22. Jensen,N.A., Pedersen,K.M., Celis,J.E., & West,M.J.
Failure of central nervous system myelination in MBP/c-myc transgenic
mice: evidence for c-myc cytotoxicity. Oncogene.16, 2123-2129
(1998).
23. Proepper,C. et al. Abelson interacting
protein 1 (Abi-1) is essential for dendrite morphogenesis and synapse
formation. EMBO J.26, 1397-1409 (2007).
24. Benoist,M.,
Gaillard,S., & Castets,F. The striatin family: a new signaling platform
in dendritic spines. J. Physiol Paris.99, 146-153 (2006).
25. Tuckova,L. et al. Anti-gliadin
antibodies in patients with celiac disease cross-react with enterocytes
and human calreticulin. Clin. Immunol. Immunopathol.85,
289-296 (1997).
26. Alaedini,A. et al. Immune cross-reactivity
in celiac disease: anti-gliadin antibodies bind to neuronal synapsin
I. J. Immunol.178, 6590-6595 (2007).
27. Agarwal,A., Sarkar,S., Nazabal,C., Balasundaram,G., & Rao,K.V.
B cell responses to a peptide epitope. I. The cellular basis for restricted
recognition. J. Immunol.157, 2779-2788 (1996).
28. Marsh,M.N. Gluten, major histocompatibility
complex, and the small intestine. A molecular and immunobiologic approach
to the spectrum of gluten sensitivity ('celiac sprue'). Gastroenterology.102,
330-354 (1992).
29. Garey,L.J. et al. Reduced dendritic
spine density on cerebral cortical pyramidal neurons in schizophrenia. J.
Neurol. Neurosurg. Psychiatry.65, 446-453 (1998).
30. Uranova,N.A. et al. The role of
oligodendrocyte pathology in schizophrenia. Int. J. Neuropsychopharmacol.10,
537-545 (2007).
31. Uranova,N.A., Vostrikov,V.M., Orlovskaya,D.D., & Rachmanova,V.I.
Oligodendroglial density in the prefrontal cortex in schizophrenia and
mood disorders: a study from the Stanley Neuropathology Consortium. Schizophr.
Res.67, 269-275 (2004).
32. Buonanno,A. et al. Neuregulins
and neuronal plasticity: possible relevance in schizophrenia. Novartis.
Found. Symp.289, 165-177 (2008).
33. Seeman,P. Glutamate and dopamine components
in schizophrenia. J. Psychiatry Neurosci.34, 143-149 (2009).
34. Hennah,W. & Porteous,D. The DISC1
pathway modulates expression of neurodevelopmental, synaptogenic and
sensory perception genes. PLoS. One.4, e4906 (2009).
35. Pardridge,W.M. Re-engineering biopharmaceuticals
for delivery to brain with molecular Trojan horses. Bioconjug. Chem.19,
1327-1338 (2008).
37. Patterson,P.H. Immune involvement in schizophrenia
and autism: etiology, pathology and animal models. Behav. Brain Res.204,
313-321 (2009).
38. Rothermundt,M., Arolt,V., & Bayer,T.A.
Review of immunological and immunopathological findings in schizophrenia. Brain
Behav. Immun.15, 319-339 (2001).
39. Ganguli,R., Rabin,B.S., Kelly,R.H., Lyte,M., & Ragu,U.
Clinical and laboratory evidence of autoimmunity in acute schizophrenia. Ann. N. Y. Acad. Sci.496, 676-685 (1987).
40. O'Donnell,M.C. et al.Increased production
of interleukin-2 (IL-2) but not soluble interleukin-2 receptors (sIL-2R)
in unmedicated patients with schizophrenia and schizophreniform disorder. Psychiatry
Res.65, 171-178 (1996).
41. Thompson,P.M. et al. Mapping cortical
change in Alzheimer's disease, brain development, and schizophrenia. Neuroimage.23
Suppl 1, S2-18 (2004).
42. Huebner,F.R., Lieberman,K.W., Rubino,R.P., & Wall,J.S.
Demonstration of high opioid-like activity in isolated peptides from
wheat gluten hydrolysates. Peptides.5, 1139-1147 (1984).
43. Laparra Llopis,J.M. & Sanz,H.Y. Gliadins
induce TNFalpha production through cAMP-dependent protein kinase A activation
in intestinal cells (Caco-2). J. Physiol Biochem.66, 153-159
(2010).
44. Lammers,K.M. et al. Gliadin induces
an increase in intestinal permeability and zonulin release by binding
to the chemokine receptor CXCR3. Gastroenterology.135,
194-204 (2008).
45. Barone,M.V. et al. Growth factor-like
activity of gliadin, an alimentary protein: implications for coeliac
disease. Gut.56, 480-488 (2007).
46. Jelinkova,L., Tuckova,L., Cinova,J., Flegelova,Z., & Tlaskalova-Hogenova,H.
Gliadin stimulates human monocytes to production of IL-8 and TNF-alpha
through a mechanism involving NF-kappaB. FEBS Lett.571,
81-85 (2004).
47. Carter,C.J. Alzheimer's disease: A pathogenetic
autoimmune disorder caused by Herpes simplex in a gene-dependent manner. Int.
J. Alz. Disin press, (2010).
48. Kanduc,D. Describing the hexapeptide identity
platform between the influenza A H5N1 and Homo sapiens proteomes. Biologics.4,
245-261 (2010).
49. Katzourakis,A. & Gifford,R.J. Endogenous
Viral Elements in Animal Genomes. Plos. Genet.6, e1001191.
doi:10.1371/journal.pgen.1001191 (2010).
50. Brocchieri,L. & Karlin,S. Protein
length in eukaryotic and prokaryotic proteomes. Nucleic Acids Res.33,
3390-3400 (2005).
51. Jia,P., Wang,L., Meltzer,H.Y., & Zhao,Z.
Common variants conferring risk of schizophrenia: a pathway analysis
of GWAS data. Schizophr. Res.122, 38-42 (2010).
52. Altschul,S.F. et al. Gapped BLAST
and PSI-BLAST: a new generation of protein database search programs. Nucleic
Acids Res.25, 3389-3402 (1997).
53. Larsen,J.E., Lund,O., & Nielsen,M.
Improved method for predicting linear B-cell epitopes. Immunome.
Res.2, 2 (2006).
54. Bendel,O., Meijer,B., Hurd,Y., & von
Euler,G. Cloning and expression of the human NMDA receptor subunit NR3B
in the adult human hippocampus. Neurosci. Lett.377, 31-36
(2005).
55. Sakagami,H. et al. IQ-ArfGEF/BRAG1
is a guanine nucleotide exchange factor for Arf6 that interacts with
PSD-95 at postsynaptic density of excitatory synapses. Neurosci. Res.60,
199-212 (2008).
56. Jaffrey,S.R., Benfenati,F., Snowman,A.M.,
Czernik,A.J., & Snyder,S.H. Neuronal nitric-oxide synthase localization
mediated by a ternary complex with synapsin and CAPON. Proc. Natl.
Acad. Sci. U. S. A.99, 3199-3204 (2002).
57. Carrel,D. et al. NOS1AP regulates
dendrite patterning of hippocampal neurons through a carboxypeptidase
E-mediated pathway. J. Neurosci.29, 8248-8258 (2009).
58. Schoch,S. et al. Redundant functions
of RIM1alpha and RIM2alpha in Ca(2+)-triggered neurotransmitter release. EMBO
J.25, 5852-5863 (2006).
59. Hosaka,M. & Sudhof,T.C. Synapsin III,
a novel synapsin with an unusual regulation by Ca2+. J. Biol. Chem.273,
13371-13374 (1998).
60. Ye,B. et al. GRASP-1: a neuronal
RasGEF associated with the AMPA receptor/GRIP complex. Neuron.26,
603-617 (2000).
61. Szebenyi,G. et al. Activity-driven
dendritic remodeling requires microtubule-associated protein 1A. Curr.
Biol.15, 1820-1826 (2005).
62. Bockers,T.M. et al. Synaptic scaffolding
proteins in rat brain. Ankyrin repeats of the multidomain Shank protein
family interact with the cytoskeletal protein alpha-fodrin. J. Biol.
Chem.276, 40104-40112 (2001).
63. Eastwood,S.L. et al. Altered expression
of synaptic protein mRNAs in STOP (MAP6) mutant mice. J. Psychopharmacol.21,
635-644 (2007).
64. Mason,J.L. et al. ATF5 regulates
the proliferation and differentiation of oligodendrocytes. Mol. Cell
Neurosci.29, 372-380 (2005).
66. Liu,A., Zhuang,Z., Hoffman,P.W., & Bai,G.
Functional analysis of the rat N-methyl-D-aspartate receptor 2A promoter:
multiple transcription starts points, positive regulation by Sp factors,
and translational regulation. J. Biol. Chem.278, 26423-26434
(2003).
67. Ma,L., Song,L., Radoi,G.E., & Harrison,N.L.
Transcriptional regulation of the mouse gene encoding the alpha-4 subunit
of the GABAA receptor. J. Biol. Chem.279, 40451-40461
(2004).
69. Enard,W. et al. A humanized version
of Foxp2 affects cortico-basal ganglia circuits in mice. Cell.137,
961-971 (2009).
70. Hisaoka,T., Nakamura,Y., Senba,E., & Morikawa,Y.
The forkhead transcription factors, Foxp1 and Foxp2, identify different
subpopulations of projection neurons in the mouse cerebral cortex. Neuroscience.166,
551-563 (2010).
71. Konopka,G. et al. Human-specific
transcriptional regulation of CNS development genes by FOXP2. Nature.462,
213-217 (2009).
72. Groszer,M. et al. Impaired synaptic
plasticity and motor learning in mice with a point mutation implicated
in human speech deficits. Curr. Biol.18, 354-362 (2008).
73. Nakatani,T., Kumai,M., Mizuhara,E., Minaki,Y., & Ono,Y.
Lmx1a and Lmx1b cooperate with Foxa2 to coordinate the specification
of dopaminergic neurons and control of floor plate cell differentiation
in the developing mesencephalon. Dev. Biol.339, 101-113
(2010).
74. Ji,H. et al. Tuning the excitability
of midbrain dopamine neurons by modulating the Ca2+ sensitivity of SK
channels. Eur. J. Neurosci.29, 1883-1895 (2009).
75. Jacobsen,J.P. et al. SK3 K+ channel-deficient
mice have enhanced dopamine and serotonin release and altered emotional
behaviors. Genes Brain Behav.7, 836-848 (2008).
76. Lee,S. et al. Essential role for
the Prader-Willi syndrome protein necdin in axonal outgrowth. Hum.
Mol. Genet.14, 627-637 (2005).
77. Lu,B. et al. Peptide neurotransmitters
activate a cation channel complex of NALCN and UNC-80. Nature.457,
741-744 (2009).
78. Zhan,L. et al. ALG-2 interacting
protein AIP1: a novel link between D1 and D3 signalling. Eur. J. Neurosci.27,
1626-1633 (2008).
79. Scheuch,K. et al. Characterization
of a functional promoter polymorphism of the human tryptophan hydroxylase
2 gene in serotonergic raphe neurons. Biol. Psychiatry.62,
1288-1294 (2007).
80. Sunabori,T. et al. Cell-cycle-specific
nestin expression coordinates with morphological changes in embryonic
cortical neural progenitors. J. Cell Sci.121, 1204-1212
(2008).
81. Graff,J., Koshibu,K., Jouvenceau,A., Dutar,P., & Mansuy,I.M.
Protein phosphatase 1-dependent transcriptional programs for long-term
memory and plasticity. Learn. Mem.17, 355-363 (2010).
82. Xu,T.X. et al. Hyperdopaminergic
tone erodes prefrontal long-term potential via a D2 receptor-operated
protein phosphatase gate. J. Neurosci.29, 14086-14099
(2009).
83. Li,L. et al. Serotonin(1A)-receptor-dependent
signaling proteins in mouse hippocampus. Neuropharmacology.57,
556-566 (2009).
84. Kim,S.M., Ahn,S.M., Go,B.S., Wang,J.Q., & Choe,E.S.
Alterations in AMPA receptor phosphorylation in the rat striatum following
acute and repeated cocaine administration. Neuroscience.163,
618-626 (2009).
85. Kanematsu,T. et al. Modulation
of GABA(A) receptor phosphorylation and membrane trafficking by phospholipase
C-related inactive protein/protein phosphatase 1 and 2A signaling complex
underlying brain-derived neurotrophic factor-dependent regulation of
GABAergic inhibition. J. Biol. Chem.281, 22180-22189 (2006).
86. Terry-Lorenzo,R.T. et al. Neurabin/protein
phosphatase-1 complex regulates dendritic spine morphogenesis and maturation. Mol.
Biol. Cell.16, 2349-2362 (2005).
87. Rex,E.B. et al. Identification
of RanBP 9/10 as interacting partners for protein kinase C (PKC) gamma/delta
and the D1 dopamine receptor: regulation of PKC-mediated receptor phosphorylation. Mol.
Pharmacol.78, 69-80 (2010).
88. Yin,Y.X. et al. RanBPM contributes
to TrkB signaling and regulates brain-derived neurotrophic factor-induced
neuronal morphogenesis and survival. J. Neurochem.114,
110-121 (2010).
89. Bai,D., Chen,H., & Huang,B.R. RanBPM
is a novel binding protein for p75NTR. Biochem. Biophys. Res. Commun.309,
552-557 (2003).
90. Murrin,L.C. & Talbot,J.N. RanBPM,
a scaffolding protein in the immune and nervous systems. J. Neuroimmune.
Pharmacol.2, 290-295 (2007).
91. Rao,M.A. et al. RanBPM, a nuclear
protein that interacts with and regulates transcriptional activity of
androgen receptor and glucocorticoid receptor. J. Biol. Chem.277,
48020-48027 (2002).
92. Harikrishnan,K.N. et al. Alleviating
transcriptional inhibition of the norepinephrine slc6a2 transporter gene
in depolarized neurons. J. Neurosci.30, 1494-1501 (2010).
93. Chiba,H., Muramatsu,M., Nomoto,A., & Kato,H.
Two human homologues of Saccharomyces cerevisiae SWI2/SNF2 and Drosophila
brahma are transcriptional coactivators cooperating with the estrogen
receptor and the retinoic acid receptor. Nucleic Acids Res.22,
1815-1820 (1994).
94. Hsiao,P.W., Fryer,C.J., Trotter,K.W.,
Wang,W., & Archer,T.K. BAF60a mediates critical interactions between
nuclear receptors and the BRG1 chromatin-remodeling complex for transactivation. Mol.
Cell Biol.23, 6210-6220 (2003).
95. Levesque,D. & Rouillard,C. Nur77 and
retinoid X receptors: crucial factors in dopamine-related neuroadaptation. Trends
Neurosci.30, 22-30 (2007).
96. Seshadri,S. et al. Disrupted-in-Schizophrenia-1
expression is regulated by beta-site amyloid precursor protein cleaving
enzyme-1-neuregulin cascade. Proc. Natl. Acad. Sci. U. S. A.107,
5622-5627 (2010).
97. Longart,M., Liu,Y., Karavanova,I., & Buonanno,A.
Neuregulin-2 is developmentally regulated and targeted to dendrites of
central neurons. J. Comp Neurol.472, 156-172 (2004).
98. Dimayuga,F.O. et al. The neuregulin
GGF2 attenuates free radical release from activated microglial cells. J.
Neuroimmunol.136, 67-74 (2003).
99. Petersen,P.H., Zou,K., Krauss,S., & Zhong,W.
Continuing role for mouse Numb and Numbl in maintaining progenitor cells
during cortical neurogenesis. Nat. Neurosci.7, 803-811
(2004).
100. Li,H.S. et al. Inactivation of
Numb and Numblike in embryonic dorsal forebrain impairs neurogenesis
and disrupts cortical morphogenesis. Neuron.40, 1105-1118
(2003).
101. Burns,B. et al. Rai1 haploinsufficiency
causes reduced Bdnf expression resulting in hyperphagia, obesity and
altered fat distribution in mice and humans with no evidence of metabolic
syndrome. Hum. Mol. Genet.19, 4026-4042 (2010).
102. Savino,M.T. et al. Rai acts as
a negative regulator of autoimmunity by inhibiting antigen receptor signaling
and lymphocyte activation. J. Immunol.182, 301-308 (2009).
103. Darcy,M.J., Calvin,K., Cavnar,K., & Ouimet,C.C.
Regional and subcellular distribution of HDAC4 in mouse brain. J.
Comp Neurol.518, 722-740 (2010).
104. Chawla,S., Vanhoutte,P., Arnold,F.J.,
Huang,C.L., & Bading,H. Neuronal activity-dependent nucleocytoplasmic
shuttling of HDAC4 and HDAC5. J. Neurochem.85, 151-159
(2003).
105. Sugo,N. et al. Nucleocytoplasmic
translocation of HDAC9 regulates gene expression and dendritic growth
in developing cortical neurons. Eur. J. Neurosci.31, 1521-1532
(2010).
106. Hirsch,E. et al. Defective dendrite
elongation but normal fertility in mice lacking the Rho-like GTPase activator
Dbl. Mol. Cell Biol.22, 3140-3148 (2002).
107. Chen,N. & Napoli,J.L. All-trans-retinoic
acid stimulates translation and induces spine formation in hippocampal
neurons through a membrane-associated RARalpha. FASEB J.22,
236-245 (2008).
109. Ohkura,N., Hijikuro,M., & Miki,K.
Antisense oligonucleotide to NOR-1, a novel orphan nuclear receptor,
induces migration and neurite extension of cultured forebrain cells. Brain
Res. Mol. Brain Res.35, 309-313 (1996).
110. White,M.K., Johnson,E.M., & Khalili,K.
Multiple roles for Puralpha in cellular and viral regulation. Cell
Cycle.8, 1-7 (2009).
111. Khalili,K. et al. Puralpha is essential
for postnatal brain development and developmentally coupled cellular
proliferation as revealed by genetic inactivation in the mouse. Mol.
Cell Biol.23, 6857-6875 (2003).
112. Haas,S. et al. A 39-kD DNA-binding
protein from mouse brain stimulates transcription of myelin basic protein
gene in oligodendrocytic cells. J. Cell Biol.130, 1171-1179
(1995).
113. Dugas,J.C. et al. Dicer1 and miR-219
Are required for normal oligodendrocyte differentiation and myelination. Neuron.65,
597-611 (2010).
114. van der,L.S. et al. Neuroanatomical
distribution and colocalisation of nuclear receptor corepressor (N-CoR)
and silencing mediator of retinoid and thyroid receptors (SMRT) in rat
brain. Brain Res.1059, 113-121 (2005).
115. Perlmann,T. & Wallen-Mackenzie,A.
Nurr1, an orphan nuclear receptor with essential functions in developing
dopamine cells. Cell Tissue Res.318, 45-52 (2004).
116. Chen,N. & Napoli,J.L. All-trans-retinoic
acid stimulates translation and induces spine formation in hippocampal
neurons through a membrane-associated RARalpha. FASEB J.22,
236-245 (2008).
117. Jiang,P., Selvaraj,V., & Deng,W. Differentiation
of embryonic stem cells into oligodendrocyte precursors. J. Vis. Exp.1960
(2010).
118. Steplewski,A. et al. MyEF-3, a
developmentally controlled brain-derived nuclear protein which specifically
interacts with myelin basic protein proximal regulatory sequences. Biochem.
Biophys. Res. Commun.243, 295-301 (1998).
119. Cai,H. et al. The sarcoglycan complex
in Schwann cells and its role in myelin stability. Exp. Neurol.205,
257-269 (2007).
120. Chan,P. et al. Epsilon-sarcoglycan
immunoreactivity and mRNA expression in mouse brain. J. Comp Neurol.482,
50-73 (2005).
121. Waite,A., Tinsley,C.L., Locke,M., & Blake,D.J.
The neurobiology of the dystrophin-associated glycoprotein complex. Ann.
Med.41, 344-359 (2009).
122. Goryunov,D., He,C.Z., Lin,C.S., Leung,C.L., & Liem,R.K.
Nervous-tissue-specific elimination of microtubule-actin crosslinking
factor 1a results in multiple developmental defects in the mouse brain. Mol.
Cell Neurosci.44, 1-14 (2010).
123. Sanchez-Soriano,N. et al. Mouse
ACF7 and drosophila short stop modulate filopodia formation and microtubule
organisation during neuronal growth. J. Cell Sci.122,
2534-2542 (2009).
124. Ge,X., Frank,C.L., Calderon,d.A., & Tsai,L.H.
Hook3 interacts with PCM1 to regulate pericentriolar material assembly
and the timing of neurogenesis. Neuron.65, 191-203 (2010).
125. Buchman,J.J. et al. Cdk5rap2 interacts
with pericentrin to maintain the neural progenitor pool in the developing
neocortex. Neuron.66, 386-402 (2010).
126. Poirier,K. et al. Mutations in
the neuronal ss-tubulin subunit TUBB3 result in malformation of cortical
development and neuronal migration defects. Hum. Mol. Genet.19,
4462-4473 (2010).
127. Chidambaram,S., Zimmermann,J., & von
Mollard,G.F. ENTH domain proteins are cargo adaptors for multiple SNARE
proteins at the TGN endosome. J. Cell Sci.121, 329-338
(2008).
128. Xie,L.X., Calafat,J., Janssen,H., de,l.,
I, & Mollinedo,F. Intracellular location of syntaxin 7 in human neutrophils. Immunol.
Lett.129, 72-77 (2010).
129. Achuthan,A. et al. Regulation of
the endosomal SNARE protein syntaxin 7 by colony-stimulating factor 1
in macrophages. Mol. Cell Biol.28, 6149-6159 (2008).
130. Collins,R.F., Schreiber,A.D., Grinstein,S., & Trimble,W.S.
Syntaxins 13 and 7 function at distinct steps during phagocytosis. J.
Immunol.169, 3250-3256 (2002).
131. Mullock,B.M. et al. Syntaxin 7
is localized to late endosome compartments, associates with Vamp 8, and
Is required for late endosome-lysosome fusion. Mol. Biol. Cell.11,
3137-3153 (2000).
132. Mattera,L. et al. The E1A-associated
p400 protein modulates cell fate decisions by the regulation of ROS homeostasis. Plos.
Genet.6, e1000983 (2010).
133. Lan,H.C., Li,H.J., Lin,G., Lai,P.Y., & Chung,B.C.
Cyclic AMP stimulates SF-1-dependent CYP11A1 expression through homeodomain-interacting
protein kinase 3-mediated Jun N-terminal kinase and c-Jun phosphorylation. Mol.
Cell Biol.27, 2027-2036 (2007).
134. Rochat-Steiner,V. et al. FIST/HIPK3:
a Fas/FADD-interacting serine/threonine kinase that induces FADD phosphorylation
and inhibits fas-mediated Jun NH(2)-terminal kinase activation. J.
Exp. Med.192, 1165-1174 (2000).
135. Hu,M. et al. Histone H3 lysine
36 methyltransferase Hypb/Setd2 is required for embryonic vascular remodeling. Proc.
Natl. Acad. Sci. U. S. A.107, 2956-2961 (2010).
136. Ramadan,S. et al. p73 induces apoptosis
by different mechanisms. Biochem. Biophys. Res. Commun.331,
713-717 (2005).
137. Rossi,M., Sayan,A.E., Terrinoni,A., Melino,G., & Knight,R.A.
Mechanism of induction of apoptosis by p73 and its relevance to neuroblastoma
biology. Ann. N. Y. Acad. Sci.1028, 143-149 (2004).
138. Bourdon,J.C., Renzing,J., Robertson,P.L.,
Fernandes,K.N., & Lane,D.P. Scotin, a novel p53-inducible proapoptotic
protein located in the ER and the nuclear membrane. J. Cell Biol.158,
235-246 (2002).
140. Becknell,B. et al. Hlx homeobox
transcription factor negatively regulates interferon-gamma production
in monokine-activated natural killer cells. Blood.109,
2481-2487 (2007).
141. Gemelli,C. et al. The vitamin D3/Hox-A10
pathway supports MafB function during the monocyte differentiation of
human CD34+ hemopoietic progenitors. J. Immunol.181, 5660-5672
(2008).
142. Ge,X., Frank,C.L., Calderon,d.A., & Tsai,L.H.
Hook3 interacts with PCM1 to regulate pericentriolar material assembly
and the timing of neurogenesis. Neuron.65, 191-203 (2010).
143. Ho,S.N. Intracellular water homeostasis
and the mammalian cellular osmotic stress response. J. Cell Physiol.206,
9-15 (2006).
144. Goldman,M.B. The mechanism of life-threatening
water imbalance in schizophrenia and its relationship to the underlying
psychiatric illness. Brain Res. Rev.61, 210-220 (2009).
145. Paytubi,S. et al. ABC50 promotes
translation initiation in mammalian cells. J. Biol. Chem.284,
24061-24073 (2009).
 Subscribe
in a reader
Subscribe
in a reader .
.